토머스 헌트 모건
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
토머스 헌트 모건은 1866년 켄터키주 렉싱턴에서 태어난 미국의 발생학자이자 유전학자이다. 그는 초파리를 이용한 유전 연구를 통해 유전자가 염색체 위에 존재하며, 염색체 교차 현상을 규명하여 유전학 발전에 크게 기여했다. 모건은 1933년 노벨 생리학·의학상을 수상했으며, 멘델의 유전 법칙과 염색체설을 통합하는 데 중요한 역할을 했다.
더 읽어볼만한 페이지
- 캘리포니아 공과대학교 - 고든 무어
고든 무어는 미국의 기업인이자 공학자로, 페어차일드 반도체를 거쳐 로버트 노이스와 함께 인텔을 공동 설립했으며, 집적 회로 집적도가 2년마다 2배씩 증가한다는 "무어의 법칙"으로 알려져 있고, 고든과 베티 무어 재단을 설립하여 자선 활동을 펼쳤다. - 캘리포니아 공과대학교 - 팔로마 천문대
팔로마 천문대는 캘리포니아 팔로마 산에 위치한 천문대로, 헤일 반사망원경을 포함한 다양한 장비를 통해 천문학 발전에 기여했으며 현재는 광해 문제 속에서도 연구와 대중 참여를 지속하고 있다. - 켄터키 대학교 동문 - 데빈 부커
데빈 부커는 NBA 피닉스 선즈의 슈팅 가드로, 2015년 드래프트 1라운드 13순위로 지명된 후 뛰어난 득점력과 코비 브라이언트를 연상시키는 플레이 스타일, 올-NBA 퍼스트 팀 선정, 70득점 경기 기록, 도쿄 올림픽 금메달 기여 등으로 리그 정상급 선수로 인정받고 있다. - 켄터키 대학교 동문 - 뱀 아데바요
뱀 아데바요는 나이지리아계 미국인 프로 농구 선수로, NBA 마이애미 히트에서 센터로 뛰며 뛰어난 수비와 득점력으로 팀의 핵심 선수로 성장, NBA 올스타에 세 차례 선정되었고 2020년 올림픽 농구 금메달을 획득했다. - 캘리포니아 공과대학교 교수 - 루돌프 마커스
루돌프 마커스는 캐나다 출신의 미국 화학자로서 전자 전달 이론 연구로 1992년 노벨 화학상을 수상했으며, RRKM 이론 개발에 기여했고, 여러 학술 단체 회원으로 활동하며 다수의 상을 수상한 저명한 학자이다. - 캘리포니아 공과대학교 교수 - 찰스 릭터
찰스 릭터는 미국의 지진학자로, 베노 구텐베르크와 함께 지진의 상대적 크기를 측정하는 릭터 규모를 개발했으며, 캘리포니아 공과대학교에서 지진학 교수로 재직하며 지진 연구와 교육에 헌신했다.
| 토머스 헌트 모건 - [인물]에 관한 문서 | |
|---|---|
| 기본 정보 | |
 | |
| 이름 | 토머스 헌트 모건 |
| 로마자 표기 | Teomaseu Heonteu Mogeon |
| 한자 표기 | 托馬斯·亨特·摩根 |
| 가나 표기 | トーマス・ハント・モーガン |
| 출생과 사망 | |
| 출생 | 1866년 9월 25일 |
| 출생지 | 미국1865 켄터키주 렉싱턴 |
| 사망 | 1945년 12월 4일 |
| 사망지 | 미국1912 캘리포니아주 패서디나 |
| 학문 분야 | |
| 분야 | 유전학, 발생학 |
| 연구 기관 | 브린모어 대학 컬럼비아 대학교 캘리포니아 공과대학교 우즈홀 해양 생물학 연구소 |
| 출신 학교 | 켄터키 대학교 존스 홉킨스 대학교 |
| 지도 학생 | 존 하워드 노스럽 하먼 J. 멀러 |
| 업적 | |
| 주요 업적 | 키이로쇼우조우바에 연구 유전적 연쇄 연구 |
| 수상 | 전미 과학 아카데미 회원 (1909년) 왕립 학회 외국인 회원 (1919년) 노벨 생리학·의학상 (1933년) 코플리 메달 (1939년) |
| |
2. 어린 시절과 교육
토머스 헌트 모건은 1866년 미국 켄터키주 렉싱턴에서 태어났다.[2][3] 그의 집안은 미국 남부의 유력 가문이었으며, 숙부인 존 헌트 모건은 남북전쟁 당시 남부 연합군의 장군이었다.[3] 어린 시절부터 자연사에 흥미를 보인 모건은 1886년 켄터키 주립 대학(현재의 켄터키 대학교)에서 동물학으로 이학사 학위를 수석으로 받았다.[4]
이후 존스 홉킨스 대학교 대학원에 진학하여 생물학을 전공했으며, 형태학자이자 발생학자인 윌리엄 키스 브룩스의 지도를 받았다. 그는 바다거미의 발생학 연구로 1890년 철학 박사 학위를 취득했다.[5] 박사 학위 취득 후 1년간 존스 홉킨스에 머물며 연구를 지속했고, 1891년 브린모어 대학교의 교수로 임용되었다. 대학원 시절부터 매사추세츠 주 우즈홀의 해양 생물 연구소(MBL)와 관계를 맺고 연구 활동에 참여했다.[5][6]
2. 1. 가계와 유년 시절
모건은 1866년 미국 켄터키주 렉싱턴에서 찰턴 헌트 모건과 엘렌 키 하워드 모건 사이에서 태어났다.[2][3] 그의 부계는 남부의 농장주이자 노예 소유주 가문이었으며, 숙부인 존 헌트 모건은 남북전쟁 동안 남부 연합 장군이었다.[3] 그의 증조부 존 웨슬리 헌트는 앨러게니 산맥 서부 지역 최초의 백만장자 중 한 명이었다. 모친을 통해서는, 스타 스팽글드 배너의 작가인 프랜시스 스콧 키와 메릴랜드 주 주지사이자 상원 의원이었던 존 이거 하워드의 증손자였다.[3] 남북전쟁 이후, 남부 연합을 지지했던 그의 가족은 시민권과 재산권의 일시적 상실로 어려움을 겪었다. 부친 찰턴 헌트 모건은 영사였으나 전쟁 후 정치 분야에서 일자리를 구하는 데 어려움을 겪었다.모건은 어릴 때부터 자연사에 흥미를 보였다. 16세에 예비 과정을 시작하여 켄터키 대학교(당시 켄터키 주립 대학)에 다녔으며, 과학, 특히 자연사에 집중하여 여름 동안 미국 지질 조사국에서 일하기도 했다. 그는 1886년 수석 졸업생으로 동물학 이학사 학위를 받았다.[4]
학부 졸업 후 매사추세츠 주 애니스쾀의 해양 생물 학교에서 여름을 보낸 뒤, 새로 설립된 존스 홉킨스 대학교에 들어가 생물학 대학원 과정을 밟았다. 형태학자이자 발생학자인 윌리엄 키스 브룩스 밑에서 2년간 실험 연구를 수행하며 여러 논문을 발표했고, 이를 바탕으로 1888년 켄터키 주립 대학에서 이학 석사 학위를 받았다. 그는 켄터키 주립 대학의 정교수직 제안을 받았으나 존스 홉킨스에 남아 연구를 계속했다.
브룩스의 지도 아래, 모건은 1889년과 1890년 여름 매사추세츠 주 우즈홀의 해양 생물 연구소(MBL)에서 수집한 자료를 바탕으로 바다거미의 발생학에 대한 연구를 진행했다. 그는 이 연구를 통해 바다거미가 다른 절지동물과의 계통 발생 관계에서 갑각류보다는 거미류에 더 가깝다는 결론을 내렸다. 이 연구 결과를 담은 논문으로 1890년 존스 홉킨스에서 철학 박사 학위를 받았고, 브루스 연구 펠로우십을 통해 자메이카, 바하마, 유럽 등지를 여행하며 추가 연구를 수행했다.[5] 이후 1891년 브린모어 대학교의 교수가 되었다. 그는 1890년부터 1942년까지 MBL에서 독립 연구원으로 활동했으며, 1897년부터 1945년까지는 MBL 이사로도 활동했다.[6]
2. 2. 대학교와 대학원
모건은 16세에 예비 과정을 시작으로 켄터키 주립 대학(현재의 켄터키 대학교)에 입학했다. 그는 과학, 특히 자연사에 흥미를 보였으며, 여름 동안에는 미국 지질 조사국에서 일하기도 했다. 1886년 수석 졸업생으로 이학사 학위를 받았다.[4]학부 졸업 후, 매사추세츠 주 애니스쾀의 해양 생물 학교에서 여름을 보낸 뒤, 새로 설립된 존스 홉킨스 대학교에서 동물학과 생물학 대학원 과정을 시작했다.[2][3] 존스 홉킨스에서는 형태학자이자 발생학자인 윌리엄 키스 브룩스 밑에서 2년간 실험 연구를 수행하며 여러 논문을 발표했다. 이를 바탕으로 1888년에는 모교인 켄터키 주립 대학에서 이학석사 학위를 받았다. 당시 켄터키 주립 대학은 다른 기관에서 2년간의 학습과 대학 교수진의 시험을 통과하면 석사 학위를 수여했다. 대학 측에서 정교수직을 제안했지만, 모건은 존스 홉킨스에 남아 연구를 계속하기로 결정하고 연구비를 지원받았다.
브룩스의 지도 아래, 모건은 바다거미의 발생학에 대한 연구로 박사 학위 논문을 준비했다. 이 연구는 1889년과 1890년 여름, 매사추세츠 주 우즈홀의 해양 생물 연구소(MBL)에서 수집한 자료를 바탕으로 진행되었으며, 바다거미와 다른 절지동물 간의 계통 발생 관계를 밝히는 것을 목표로 했다. 그는 연구를 통해 바다거미가 발생학적으로 갑각류보다는 거미류에 더 가깝다는 결론을 내렸다. 이 연구 결과를 발표하며 1890년 존스 홉킨스 대학교에서 철학 박사 학위를 취득했고, 브루스 연구 펠로우십(Bruce Fellowship)을 받았다. 그는 이 펠로우십을 이용해 자메이카, 바하마, 유럽 등지를 여행하며 추가 연구를 수행했다.[5] 박사 학위 취득 후 브린모어 대학교 교수가 되기 전까지 1년 동안 존스 홉킨스에 더 머물렀다.
모건은 대학원 시절부터 해양 생물 연구소(MBL)와 깊은 인연을 맺었다. 1890년부터 1942년까지 MBL에서 독립 연구원으로 활동했으며, 특히 1910년부터 1925년까지 매년 여름에는 컬럼비아 대학교의 유명한 '플라이 룸'(Fly Room) 동료들과 함께 MBL에서 연구 프로그램을 진행했다. 그는 연구 활동 외에도 1897년부터 1945년까지 MBL 이사로 활동하며 기관 운영에도 적극적으로 참여했다.[6]
3. 학문적 경력
토머스 헌트 모건은 존스 홉킨스 대학교에서 학위를 마친 후 1890년 브린모어 대학교의 부교수로 임명되어 학문적 경력을 시작했다.[7] 초기에는 발생학 연구에 집중하며, 1893년부터 발생 과정의 인과 관계를 밝히기 위해 실험적 방법을 도입했다. 1894년 나폴리에서의 연구와 한스 드리쉬와의 교류는 그가 전통적인 형태학에서 실험 발생학으로 연구 방향을 전환하는 중요한 계기가 되었다.[9] 이 시기 그는 재생과 유충 발달 등을 연구하며 '개구리 난자의 발달'(1897)과 '재생'(1901) 등의 저서를 출판했다.
초기 모건은 당시 많은 생물학자들처럼 다윈의 진화론, 특히 자연선택 메커니즘에 대해 회의적이었으며, 멘델의 이론과 염색체설에 대해서도 비판적인 입장을 보였다. 그는 이러한 이론들이 실험적으로 충분히 검증되지 않았고, 전성설과 유사한 관념적 설명을 제공한다고 생각했다.
1904년 컬럼비아 대학교의 실험동물학 교수로 자리를 옮기면서 그의 연구 중심은 점차 유전학으로 이동했다.[14] 그는 휴고 드 브리스의 돌연변이 이론을 검증하기 위해 1908년경부터 초파리(*Drosophila melanogaster*)를 이용한 실험을 시작했다. 1910년 흰 눈 돌연변이 수컷 초파리를 발견하고 그 유전 양상을 분석하여 성 연관 유전을 처음으로 밝혀냈다. 이는 유전자가 염색체 위에 존재한다는 보베리-서튼 염색체설을 강력하게 뒷받침하는 증거가 되었다.
이후 모건과 그의 연구실, 일명 '플라이 룸'(Fly Room)은 유전자 연관, 염색체 교차, 유전자 지도 작성 등 유전학의 핵심 개념들을 실험적으로 증명하고 발전시키는 데 크게 기여했다. 그의 제자 앨프리드 스터테번트는 1913년 최초의 유전자 지도를 작성했으며,[18] 1915년 모건은 스터테번트, 캘빈 브리지스, H. J. 뮬러와 함께 기념비적인 저서 멘델 유전의 메커니즘(The Mechanism of Mendelian Heredity)을 출판하여 현대 유전학의 기초를 다졌다.[20][21] 이러한 연구들을 통해 모건은 유전자가 염색체 위에 존재하는 물리적 실체임을 증명했으며, 초파리를 유전학 연구의 핵심 모델 생물로 확립했다. 한편, 그는 유전학 지식을 사회 문제에 잘못 적용하려는 우생학 운동에 대해 비판적인 입장을 견지했다.[26]
1928년 모건은 칼텍(Caltech)으로 옮겨 생물학부를 창설하고 이끌었다.[1][2] 그는 유전학, 진화, 발생학 등 다양한 분야의 연구를 장려하고 조지 비들, 테오도시우스 도브잔스키 등 우수한 연구자들을 유치하여 칼텍을 세계적인 생물학 연구 중심지로 발전시키는 데 기여했다.[2] 그는 미국 과학 아카데미 회장(1927년-1931년) 등 학계에서 활발히 활동했으며, 1933년 염색체의 유전적 기능 연구에 대한 공로로 노벨 생리학·의학상을 수상했다.[2] 1942년 은퇴 후에도 연구 활동을 계속했으며, 그의 제자 및 후학들 중 7명이 노벨상을 수상하는 등 후학 양성에도 큰 업적을 남겼다.
3. 1. 브린모어 대학교
1890년, 모건은 존스 홉킨스 대학교의 자매 학교인 브린모어 대학교(Bryn Mawr College)의 부교수 겸 생물학과장으로 임명되어, 동료 에드먼드 비처 윌슨(Edmund Beecher Wilson)의 자리를 이어받았다.[7] 그는 모든 형태학 관련 과목을 가르쳤고, 학과의 다른 교수인 자크 뢰브(Jacques Loeb)는 생리학 과목을 담당했다. 뢰브는 1년만 재직했지만, 이는 두 사람의 평생 우정의 시작이 되었다.[8] 모건은 주 5일, 하루 두 차례 강의했으며, 자신의 최신 연구 내용을 강의에 자주 포함시켰다. 열정적인 교사였지만, 그는 실험실 연구에 더 큰 관심을 보였다. 브린모어에서의 초기 몇 년 동안 그는 바다 따개비, 멍게 벌레, 개구리에 대한 기술적인 연구를 수행했다.1893년부터 모건은 발생학의 근본적인 문제를 해결하기 위해 실험적 방법을 적용하기 시작했다. 그는 발생 과정의 인과 관계를 밝히고자 초기 배아 세포인 할구에서 배아가 형성되는 과정이나 유핵난세포와 무핵난세포의 수정 문제 등을 연구했다.
1894년 모건은 1년간의 연구 휴가를 받아 나폴리의 ''Stazione Zoologica'' 실험실에서 연구를 수행했다. 이곳은 윌슨도 2년 전에 연구했던 곳이다. 그는 독일 생물학자 한스 드리쉬(Hans Driesch)와 함께 연구하며 발생에 대한 실험적 연구에 큰 자극을 받았다. 그 해 모건은 빗해파리 발생학에 대한 실험 연구를 완료하는 등 여러 프로젝트를 진행했다. 나폴리에서, 그리고 뢰브를 통해 그는 '발달 역학'(Entwicklungsmechanikde)이라는 실험 생물학 학파를 접하게 되었다. 이는 19세기 형태학에 큰 영향을 미쳤던 생명론적 자연철학(''Naturphilosophie'')에 대한 반작용으로 등장한 학파였다. 이 경험을 통해 모건은 자신의 연구 방향을 전통적인 기술(記述) 중심의 형태학에서 유기체 발달에 대한 물리적, 화학적 설명을 추구하는 실험 발생학으로 전환했다.[9]
당시 배아 발달 과정에 대한 과학적 논쟁이 활발했다. 빌헬름 루(Wilhelm Roux)의 모자이크 이론을 따르는 학자들은 유전 물질이 배아 세포들 사이에서 나뉘며, 각 세포는 성체 특정 부위로 발달하도록 예정되어 있다고 믿었다. 반면 드리쉬 등은 발생이 후성적 요인, 즉 난자의 세포질과 핵, 그리고 환경 간의 상호작용에 의해 영향을 받는다고 주장했다. 모건은 후자의 입장을 지지했다. 그는 드리쉬와의 연구를 통해 성게와 빗해파리 난자에서 분리된 초기 세포분열구가 루의 지지자들이 예측했던 것과 달리 완전한 유충으로 발달할 수 있음을 실험적으로 증명했다.[10] 이는 발생 과정에서 후성유전 및 환경적 요인의 역할을 강조하는 중요한 결과였다. 이와 관련하여 모건은 성게 난자에 염화마그네슘을 첨가하면 수정 없이도 분열을 유도할 수 있음을 보여주었다. 뢰브는 이 연구를 이어받아 인공적으로 아비 없는 개구리를 만드는 데 성공하여 유명해졌다.[11][12]
1895년 브린모어 대학교로 돌아온 모건은 정교수로 승진했다. 그의 주요 실험 연구는 재생과 유충 발달에 집중되었으며, 두 연구 모두 발생의 내부적 원인과 외부적 원인을 구별하여 루-드리쉬 논쟁을 명확히 하려는 목표를 가졌다. 그는 첫 저서인 ''The Development of the Frog's Egg영어''(개구리 난자의 발달)을 1897년에 출판했다. 또한 다양한 유기체의 재생 능력에 대한 연구를 시작하여 올챙이, 물고기, 지렁이의 접목과 재생을 연구했고, 그 결과를 모아 1901년 ''Regeneration영어''(재생)이라는 책으로 출판했다.
19세기 말 대부분의 발생학자들처럼 모건 역시 자연선택에 기반한 다윈의 진화론에 대해 회의적이었다. 그는 생물체의 복잡한 적응 현상이 우연한 작은 변이들의 축적으로 설명될 수 있다고 보지 않았으며, 다윈이 변이의 기원과 유전 메커니즘을 제대로 제시하지 못했다고 생각했다. 모건은 진화 자체는 사실로 받아들였지만, 실험적으로 검증하기 어려운 자연선택 메커니즘은 불완전하다고 여겼다. 또한 그는 멘델의 이론과 염색체설에 대해서도 비판적이었다. 이 이론들이 세포 내에 발생을 조절하는 유전 단위가 있다고 가정하는 점이, 과거의 전성설과 유사하다고 보았다. 그는 염색체가 유전 현상과 관련 있다는 점은 인정했지만, 개별 염색체가 특정 유전 형질을 운반한다는 주장에 대해서는 1909년까지 부정적인 입장을 유지했다. 멘델의 이론 역시 교배 결과를 설명하고 예측할 수는 있지만, 유전의 정확한 과정을 설명하지 못하는 순전히 가설적인 이론이라고 주장했다. 그는 염색체 쌍의 분리와 각 염색체가 다른 생식 세포로 들어간다는 사실만으로는 멘델주의자들이 주장하는 연관성을 입증하기에 증거가 불충분하다고 보았다.
1900년부터 모건은 성 결정 시스템 문제에 대한 연구를 시작했으며, 이는 네티 스티븐스(Nettie Stevens)가 Y 염색체의 역할을 발견했을 때 이전에 무시했던 주제였다. 그는 또한 자신의 초기 연구 초점이었던 진화 문제에 대한 연구도 계속했다.[13]
1904년 모건은 자신이 가르치던 브린모어 대학교 대학원생 릴리언 샘슨(Lilian Vaughan Sampson)과 결혼했다. 같은 해, 그는 컬럼비아 대학교의 실험동물학 교수직 제안을 수락하고 브린모어를 떠났다. 이후 24년간 컬럼비아 대학교에서 중요한 유전학 연구의 대부분을 수행하게 된다.
3. 2. 컬럼비아 대학교
1904년 모건은 친구이자 동료 생물학자였던 E. B. 윌슨의 초청을 받아 컬럼비아 대학교의 실험동물학 교수로 부임했다.[14] 그는 이후 1928년까지 24년 동안 이곳에 머물며 그의 가장 중요한 유전학 연구들을 수행하게 된다.컬럼비아 대학교 부임 초기, 모건은 여전히 발생학에 관심을 두면서도 점차 유전과 진화의 메커니즘 문제에 집중하기 시작했다. 그는 『진화와 적응』(Evolution and Adaptation, 1903)이라는 책을 출판했는데, 당시 많은 생물학자들처럼 생물 종들이 공통 조상으로부터 진화했다는 사실은 인정했지만, 찰스 다윈이 제시한 자연선택 메커니즘에 대해서는 회의적이었다. 모건은 생물체의 복잡한 적응 현상이 작은 변이들의 축적으로 설명되기 어렵다고 보았고, 다윈이 변이의 기원과 유전 메커니즘을 명확히 제시하지 못했다고 생각했다. 또한, 그는 당시 새롭게 재발견된 멘델의 유전법칙과 염색체가 유전 물질이라는 보베리-서튼 염색체설에 대해서도 비판적이었다. 그는 이러한 이론들이 유전자라는 가상의 단위가 발생 과정을 미리 결정한다고 가정하는 점에서, 과거의 전성설과 유사하다고 보았다. 그는 염색체가 유전 현상과 관련 있다는 점은 인정했지만, 개별 염색체가 특정 형질을 결정한다는 생각에는 동의하지 않았다.[15]

이러한 배경 속에서 모건은 휴고 드 브리스가 제시한 돌연변이 이론(진화가 갑작스러운 큰 변이, 즉 돌연변이를 통해 일어난다는 이론)을 실험적으로 검증하고자 했다. 1908년경, 그는 찰스 W. 우드워스와 윌리엄 E. 캐슬 등의 선행 연구를 따라 실험 재료로 초파리(*Drosophila melanogaster*)를 선택하고 연구를 시작했다. 학생 페르난두스 페인과 함께 물리적, 화학적, 방사선 등 다양한 방법으로 초파리에게 인위적인 돌연변이를 유도하려 했으나,[17][16] 처음 2년 동안은 뚜렷한 유전적 변이를 발견하지 못했다.[17]

그러던 1910년, 모건은 자신이 기르던 수많은 붉은 눈의 정상 초파리들 사이에서 우연히 흰 눈을 가진 돌연변이 수컷 한 마리를 발견했다. 그는 이 흰 눈 수컷을 정상적인 붉은 눈 암컷과 교배시켰다. 그 결과 태어난 1세대(F1) 자손들은 모두 붉은 눈을 가졌다. 다시 이 F1 세대끼리 교배시키자, 2세대(F2)에서는 붉은 눈 초파리와 흰 눈 초파리가 일정한 비율로 나타났는데, 놀랍게도 흰 눈을 가진 초파리는 모두 수컷이었다. 이 결과를 설명하기 위해 모건은 흰 눈 형질이 성염색체 중 하나인 X염색체를 통해 유전되는 성 연관 유전이라는 가설을 세웠다. 즉, 흰 눈 유전자는 X염색체 위에 있으며, 수컷(XY)은 X염색체를 하나만 가지므로 이 유전자가 발현되면 바로 흰 눈이 되지만, 암컷(XX)은 X염색체를 두 개 가지므로 두 염색체 모두에 흰 눈 유전자가 있어야만 흰 눈이 된다는 것이다. 이는 멘델의 유전법칙과 염색체설을 연결하는 중요한 발견이었다.
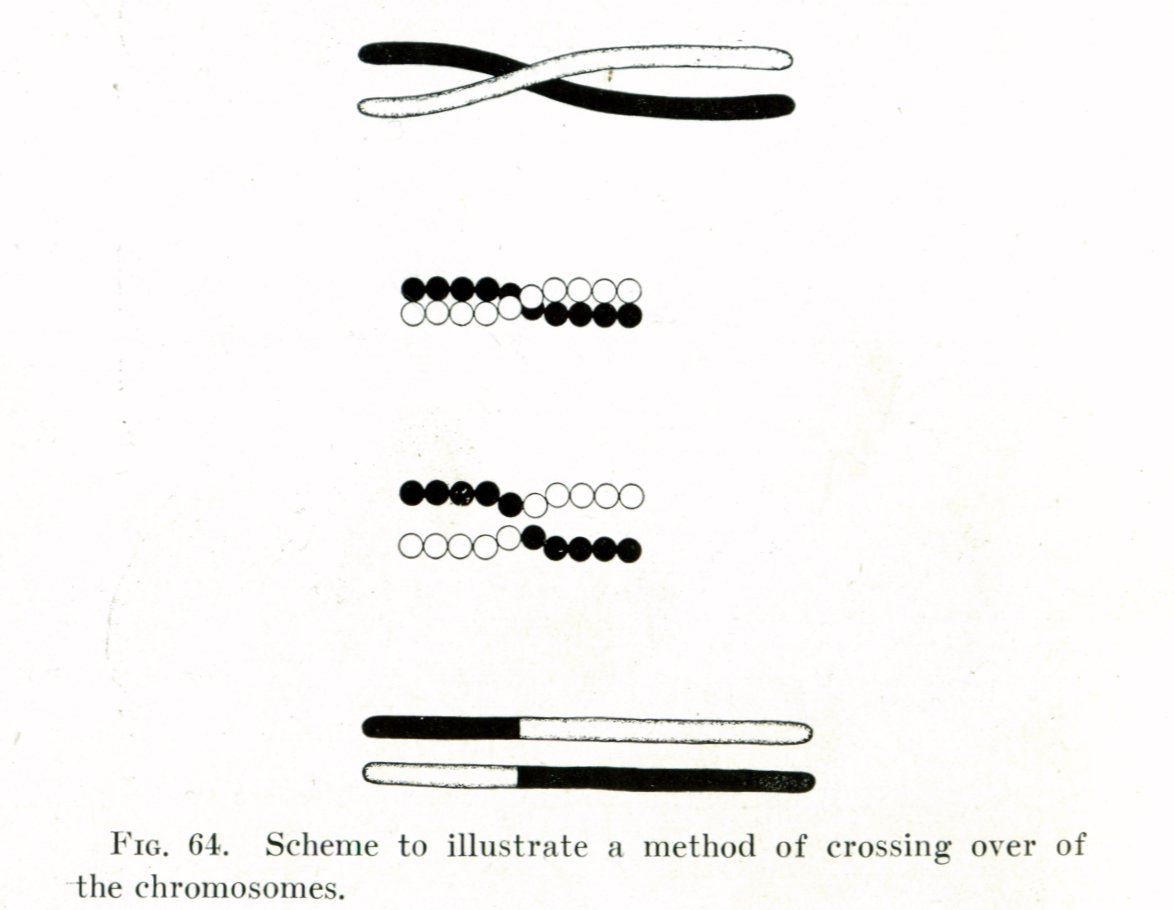
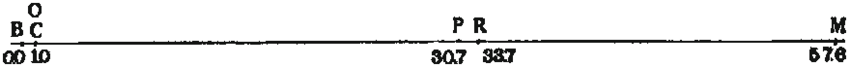
이후 모건과 그의 제자들은 더 많은 초파리 돌연변이들을 발견하고 연구했다. 특히, 여러 유전 형질들이 함께 유전되는 경향(유전자 연관)을 관찰하면서, 같은 염색체 위에 있는 유전자들이 함께 전달된다는 사실을 확인했다. 하지만 때로는 연관된 형질들이 분리되어 나타나는 현상도 발견했는데, 이는 1909년 벨기에의 프란스 알폰스 얀센스가 제안했던 염색체 간의 유전 물질 교환, 즉 염색체 교차(crossing over) 때문이라고 설명했다. 모건은 교차가 일어나는 빈도가 유전자들 사이의 거리에 비례할 것이라고 추론했고, 이를 이용하면 염색체 상에서 유전자들의 상대적인 위치를 나타내는 유전자 지도를 만들 수 있다고 생각했다. 1913년, 모건의 제자였던 앨프리드 스터테번트는 이 아이디어를 바탕으로 최초의 유전자 지도를 작성했다.[18]

모건의 연구실, 일명 '플라이 룸'(Fly Room)은 앨프리드 스터테번트, 캘빈 브리지스, H. J. 뮬러와 같은 뛰어난 젊은 연구자들과 함께 활발한 연구를 수행하며 세계적인 명성을 얻었다. 1915년, 이들 네 사람은 초파리 연구 결과를 집대성한 기념비적인 저서 『멘델 유전의 메커니즘』(The Mechanism of Mendelian Heredity)을 출판했다.[20] 이 책은 현대 유전학의 기초를 다진 교과서로 평가받는다.[21] 모건과 그의 그룹은 유전자가 염색체 위에 일직선으로 배열되어 있으며, 이들의 물리적 위치가 유전자 지도상의 위치와 일치한다는 것을 실험적으로 증명함으로써, 유전자가 더 이상 추상적인 개념이 아니라 염색체라는 물질적 실체 위에 존재함을 명확히 밝혔다. 이로써 월터 서튼과 테오도어 보베리가 제안했던 보베리-서튼 염색체설은 확고한 실험적 증거를 얻게 되었고, 1915년 이후 대부분의 생물학자들에게 받아들여졌다. 과거 멘델 법칙과 염색체설에 회의적이었던 모건 자신도 이러한 실험 결과들을 통해 자신의 입장을 바꾸게 되었다.
모건의 연구는 유전학 발전에 결정적인 영향을 미쳤다. 그의 연구 이전까지 유전자는 추상적인 개념에 가까웠으나, 그의 연구를 통해 유전자는 염색체 위에 위치하는 구체적인 물질적 단위로 인식되기 시작했다. 또한, 그는 초파리를 유전학 연구의 강력한 모델 생물로 확립시켰다. 그의 연구실은 전 세계 유전학 연구의 중심지가 되었고, 초파리는 여러 연구실에서 널리 사용되는 실험 동물이 되었다.[24] 모건은 연구가 진행됨에 따라 점차 제자들에게 연구의 주도권을 넘겨주었으며, 자신은 다시 발생학 연구에 관심을 기울이기도 했다.[25] 한편, 그는 당시 우생학 운동이 유전학적 지식을 인종 차별적인 방식으로 왜곡하여 적용하는 것에 대해 강력히 비판했다.[26]
1927년, 컬럼비아 대학교에서 25년간의 연구 활동을 마무리할 무렵, 모건은 조지 엘러리 헤일의 제안을 받아들여 캘리포니아 공과대학교(Caltech)에 새로운 생물학부를 설립하기 위해 서부로 떠나기로 결정했다. 그의 '플라이 룸' 출신 제자 및 후학들 중 7명이 훗날 노벨상을 수상하며 그의 학문적 영향력을 입증했다.
3. 3. 캘리포니아 공과대학교
1928년 모건은 칼텍(Caltech)으로 자리를 옮겨 생물학부 창설을 주도하고 부서장을 맡았다.[1][2] 그는 칼텍 생물학부를 설립하며, 존스 홉킨스 대학교나 컬럼비아 대학교와는 차별화된 연구 프로그램을 구상하여 유전학, 진화, 실험 발생학, 생리학, 생물 물리학, 생화학 연구에 중점을 두었다.[2] 또한 코로나 델 마르에 해양 연구소 설립에도 기여했다.[1][2]
모건은 칼텍 생물학부에 최고의 인재들을 유치하고자 노력했다. 컬럼비아 대학교 시절 동료였던 스터티번트, 브리지스, 잭 슐츠, 앨버트 타일러를 영입했고, 테오도시우스 도브잔스키를 국제 연구 펠로우로 초빙했다.[2] 그의 명성에 이끌려 조지 비들, 보리스 에프루시, 에드워드 L. 테이텀, 라이너스 폴링, 프리츠 벤트, 에드워드 B. 루이스, 시드니 W. 바이언스 등 많은 저명한 과학자들이 칼텍 생물학부에 합류했다.[2]
모건은 미국 과학계에서도 활발히 활동했다. 1927년부터 1931년까지 미국 과학 아카데미 회장을 지냈고, 1930년에는 미국 과학 진흥 협회 회장을 역임했다.[2] 1932년에는 뉴욕주 이타카에서 개최된 제6회 국제 유전학 회의 의장을 맡았다.[2]
1933년 모건은 염색체의 유전적 역할을 규명한 공로로 노벨 생리학·의학상을 수상했다. 그는 이미 1919년과 1930년에도 같은 업적으로 노벨상 후보로 추천된 바 있다.[2] 모건은 연구 성과가 동료들과의 협력 덕분임을 강조하며, 노벨상 상금을 브리지스, 스터트번트, 그리고 자신의 자녀들과 나누었다.[2] 그는 1933년 시상식에는 불참하고 이듬해인 1934년에 참석했는데, 이는 1933년 초파리 침샘에서 거대 다사 염색체가 재발견되어 연구 결과를 시각적으로 확인할 수 있게 된 점이 영향을 미쳤을 가능성이 있다.[2] 모건은 노벨상 수상 연설에서 유전 상담 외에 유전학이 의학 발전에 기여할 수 있는 부분을 다소 제한적으로 평가하기도 했다.[2] 1939년에는 영국 왕립 학회로부터 코플리 메달을 수여받았다.[2]
모건은 칼텍에서 두 차례 계약을 연장한 후 1942년에 은퇴하여 명예 교수이자 명예 학과장이 되었다. 그의 재직 기간은 총 14년이었다.[2] 1946년 조지 비들이 그의 후임으로 칼텍 생물학부 학과장으로 부임했다.[2] 은퇴 후에도 모건은 연구실을 유지하며 실험 연구를 지속했고, 초기 관심 분야였던 성 분화, 재생, 발생학 문제로 돌아가 연구를 이어갔다.[1][2] 그는 유전학과 발생학 사이의 이론적 연관성을 인지했지만, 이를 명확히 규명하고 실험적으로 증명하는 데에는 어려움을 느꼈다.[1]
4. 초파리 연구와 유전학의 발전
1908년경부터 컬럼비아 대학교에서 초파리(''Drosophila melanogaster'')를 이용한 유전 연구에 착수하면서 현대 유전학 발전에 결정적인 기여를 했다. 당시 멘델의 유전법칙이 재발견된 지 얼마 되지 않았고 유전자의 실체나 염색체와의 관계가 불분명한 상황에서, 모건은 실험적 접근을 통해 유전의 메커니즘을 밝히고자 했다. 그는 초기에 다윈의 자연 선택 이론이나 멘델의 유전법칙, 염색체설에 대해 회의적인 입장이었으나, 자신의 연구 결과를 통해 점차 이를 수용하고 발전시켰다.
모건 연구의 핵심적인 돌파구는 1910년 흰 눈 돌연변이 초파리를 발견하면서 마련되었다. 이 돌연변이의 유전 양상을 분석하여 특정 형질이 성별과 연관되어 유전되는 성 연관 유전을 세계 최초로 증명했으며, 이는 유전자가 X염색체 위에 존재한다는 강력한 증거가 되었다. 이 발견은 유전자가 염색체라는 물리적 구조물 위에 위치한다는 염색체설을 결정적으로 뒷받침했다.
이후 모건과 그의 연구팀, 일명 '플라이 룸(Fly Room)'의 동료들(앨프리드 스터티번트, 캘빈 브리지스, H. J. 멀러 등)은 다양한 초파리 돌연변이를 이용하여 유전자 연관과 염색체 교차 현상을 발견했다. 같은 염색체 상의 유전자들이 함께 유전되는 경향(연관)과 이 연관이 교차를 통해 깨질 수 있다는 사실을 밝혀냈으며, 교차 빈도를 이용하여 유전자 간의 상대적 거리를 추정할 수 있다는 아이디어를 제시했다. 이를 바탕으로 그의 제자 스터티번트는 1913년 세계 최초의 유전자 지도를 작성했다.[18]
1915년 모건과 그의 핵심 제자들은 이러한 연구 결과를 집대성한 기념비적인 저서 The Mechanism of Mendelian Heredity|멘델 유전의 메커니즘eng을 출판했다.[20] 이 책은 유전자가 염색체 위에 선상으로 배열되어 있으며, 멘델의 법칙이 염색체의 행동으로 설명될 수 있음을 명확히 제시하며 현대 유전학의 기초를 확립했다는 평가를 받는다.[21]
모건의 연구는 추상적인 개념이었던 유전자를 염색체 위의 구체적인 물리적 단위로 자리매김하게 했으며, 초파리를 유전학 연구의 핵심적인 모델 생물로 확립시키는 계기가 되었다.[24] 또한, 그는 당시 성장하던 우생학 운동과 그것이 인종 차별적 견해를 정당화하는 방식에 대해 비판적인 입장을 견지하기도 했다.[26]
1928년 모건은 캘리포니아 공과대학교로 자리를 옮겨 생물학과 창설을 주도했으며, 이후에도 유전학 연구를 지속하는 한편, 초기 관심사였던 실험발생학 연구로 돌아가 유전과 발생 사이의 연관성을 탐구하고자 노력했다.
4. 1. 초파리 연구의 시작
모건은 1908년경부터 초파리 ''Drosophila melanogaster''를 이용한 유전 연구를 시작했다. 당시 그는 다윈의 자연 선택 이론과 라마르크주의의 획득 형질의 유전 이론에 대해 회의적이었으며, 휴고 드 브리스가 제안한 돌연변이 이론을 실험적으로 검증하고자 했다. 또한 멘델의 유전 법칙과 염색체설에 대해서도 초기에는 확신을 갖지 못했다.찰스 W. 우드워스와 윌리엄 E. 캐슬 등의 영향을 받아 초파리 연구에 착수했으며, 페르난두스 페인과 같은 학생들과 함께 물리적, 화학적, 방사선적 방법으로 돌연변이를 유도하려 시도했으나[17][16], 약 2년간 뚜렷한 유전적 변이를 발견하지 못했다.[17]
마침내 1910년, 모건은 자신이 기르던 붉은 눈의 야생형 초파리 집단 중에서 흰 눈을 가진 돌연변이 수컷 한 마리를 발견했다. 이 흰 눈 수컷을 정상적인 붉은 눈 암컷과 교배시키자 제1대(F1) 자손은 모두 붉은 눈을 가졌다. 그러나 F1 세대끼리 교배시켜 얻은 제2대(F2)에서는 일부 흰 눈 초파리가 나타났는데, 이들은 예외 없이 모두 수컷이었다.
이 결과를 설명하기 위해 모건은 흰 눈 형질이 성 연관 유전이며, X염색체를 통해 전달된다는 가설을 세웠다. 그는 이 유전자를 '백색'(white)이라고 명명했다. 이후 다른 반성 형질들도 연이어 발견되었고, 이들이 함께 유전되는 경향을 보이자 모건은 X염색체가 여러 유전 단위를 가지고 있다고 확신하게 되었다. 그는 덴마크의 식물학자 빌헬름 요한센이 1909년에 도입한 유전자 개념을 채택하고, 유전자들이 염색체 위에 일직선으로 배열되어 있을 것이라고 결론지었다. 이러한 교배 실험 결과와 세포학적 증거들은 멘델 이론과 염색체설을 동시에 뒷받침하는 강력한 증거가 되었고, 모건은 이 두 이론에 대한 회의적인 시각을 버리고 이를 통합적으로 수용하게 되었다.
컬럼비아 대학교의 제자들인 앨프리드 스터티번트, 캘빈 브리지스, H. J. 멀러 등과의 공동 연구를 통해 모건은 초파리 연구를 포괄적인 유전 이론으로 발전시켰다. 특히, 같은 염색체 상에 있는 유전자들이 함께 유전되는 경향(유전자 연관)과 때로는 연관이 깨지는 현상(염색체 교차)을 발견하고 설명했다. 교차 현상은 1909년 벨기에 루뱅 가톨릭 대학교의 프란스 알폰스 얀센스가 관찰하고 '키아스마티피'라고 명명한 염색체의 교차 구조 관찰에 이론적 기반을 두었다. 모건은 연관된 유전자 사이의 교차 빈도가 염색체 상에서 유전자 간의 상대적 거리를 나타낼 수 있다고 제안했다.
이 아이디어를 바탕으로 모건의 제자 앨프리드 스터티번트는 1913년 세계 최초로 유전자 지도를 작성했다.[18] 1915년 모건, 스터티번트, 브리지스, 멀러는 이러한 초기 연구 결과들을 집대성한 기념비적인 저서 The Mechanism of Mendelian Heredity|멘델 유전의 메커니즘eng을 출판했다.[20] 이 책은 현대 유전학의 기초를 다진 교과서로 평가받는다.[21]
모건의 연구는 이전까지 추상적인 개념으로 여겨졌던 유전자가 염색체 위에 실제로 존재하는 물리적 실체임을 증명했으며, 초파리를 유전학 연구의 핵심적인 모델 생물로 확립하는 데 결정적인 기여를 했다. 그의 연구실은 '파리 방'(Fly Room)으로 불리며 세계적인 명성을 얻었다.
4. 2. 유전자 연관과 교차
1908년부터 초파리(''Drosophila melanogaster'')를 이용한 교배 연구를 시작했다. 1909년, 그는 자신이 기르던 초파리 중에서 눈이 흰 돌연변이 수컷 한 마리를 발견했다. 이는 일반적인 붉은 눈의 야생형과는 다른 특징이었다. 모건은 이 흰눈 수컷을 정상적인 붉은 눈 암컷과 교배시켰고, 그 결과 태어난 1세대(F1) 자손들은 모두 붉은 눈을 가졌다. 다시 F1 세대끼리 교배시키자, 2세대(F2)에서는 일부 흰눈 초파리가 나타났는데, 이들은 모두 수컷이었다.[17][16]이 현상을 설명하고자 모건은 특정 형질이 성별과 연관되어 유전된다는 성 연관 유전 가설을 세웠다. 그는 흰눈 형질이 암컷의 X염색체와 관련이 있을 것으로 추정했다. 이후 다른 여러 유전적 변이들도 성 연관 유전된다는 사실이 밝혀지면서, 모건은 X염색체가 여러 유전 단위를 가지고 있다고 확신하게 되었다. 그는 덴마크의 식물학자 빌헬름 요한센이 1909년에 도입한 유전자 개념을 받아들여, 유전자가 염색체 위에 일직선으로 배열되어 있을 것이라고 결론지었다. 교배 실험 결과와 세포학적 증거들이 멘델의 유전법칙과 염색체설을 모두 뒷받침한다는 것을 확인한 모건은 이 이론들에 대한 초기 회의적인 입장을 버리고 이를 적극적으로 수용했다.
컬럼비아 대학교에서 모건과 함께 연구한 앨프리드 스터티번트, 캘빈 브리지스, 헤르만 조지프 멀러 등은 초파리 연구를 통해 포괄적인 유전 이론을 발전시키는 데 기여했다. 특히 중요한 성과는 유전자가 염색체 상의 특정 위치에 존재하며, 이 위치를 나타내는 유전자 지도를 만들 수 있다는 것을 보여준 것이다.
모건은 성 염색체에 위치하면서도 때때로 흰눈 돌연변이와 독립적으로 유전되는 것처럼 보이는 소형 날개 돌연변이를 관찰하면서 유전자 연관이라는 개념에 도달했다. 이는 특정 유전자들이 같은 염색체 위에 존재하여 함께 유전되는 경향이 있음을 의미한다. 그러나 연관된 유전자들이 항상 함께 유전되는 것은 아니었는데, 모건은 이를 설명하기 위해 염색체 교차 현상을 가정했다. 이는 1909년 벨기에 루뱅 대학교의 프란스 알폰스 얀센스 교수가 발견하고 '키아스마티피'라고 명명한 현상에 기반한 것이었다. 모건은 연관된 유전자들 사이의 교차 빈도가 다르며, 이 빈도가 염색체 상에서 유전자들이 떨어진 거리를 나타낼 수 있다고 제안했다. 즉, 두 유전자 사이의 거리가 멀수록 교차가 일어날 확률이 높아진다는 것이다. 이 아이디어를 바탕으로 모건의 제자 앨프리드 스터티번트는 1913년 최초의 유전자 지도를 개발했다.[18] 유전자 간의 상대적 거리를 나타내는 단위는 훗날 J. B. S. 홀데인에 의해 모건의 이름을 따 센티모건(cM)으로 명명되었다.
모건은 세포학적 연구를 통해 유전자 지도상의 위치가 염색체 상의 실제 위치와 일치함을 보여주었고, 이는 멘델이 제안한 유전 요인이 염색체라는 물리적 구조에 기반한다는 결정적인 증거가 되었다. 1915년, 모건, 스터티번트, 브리지스, 멀러는 이러한 연구 결과를 종합하여 획기적인 저서인 《멘델 유전의 메커니즘》(The Mechanism of Mendelian Heredity)을 출판했다.[20] 이 책은 현대 유전학의 기초를 다진 중요한 교과서로 평가받는다.[21]
모건과 그의 동료들의 연구는 월터 서튼과 테오도어 보베리가 1902-1903년에 독립적으로 제안했던 보베리-서튼 염색체설을 확립하는 데 결정적인 역할을 했다. 이들의 연구를 통해 유전자가 염색체 위에 존재하며, 염색체의 행동을 통해 유전 현상을 설명할 수 있다는 사실이 널리 받아들여지게 되었다. 모건의 연구실, 일명 '플라이 룸'(Fly Room)은 세계적인 유전학 연구의 중심지가 되었으며, 초파리는 유전학 연구의 핵심적인 모델 생물로 자리 잡았다.[24]
4. 3. 유전자 지도 작성
모건은 많은 반성 형질들이 함께 유전된다는 사실을 통해 X염색체가 여러 유전 단위를 지니고 있음을 확신하게 되었다. 그는 덴마크의 식물학자 빌헬름 요한센이 1909년 도입한 유전자 개념을 받아들여, 유전자가 염색체 위에 일직선으로 배열되어 있을 것이라고 결론지었다.컬럼비아 대학교에서 연구를 진행하며, 모건은 성 염색체에 위치하면서도 때때로 독립적으로 분리되는 돌연변이(예: 백안 유전자와 소형 날개 유전자)를 관찰하였다. 이는 유전자 연관과 염색체 교차(crossing over) 현상에 대한 아이디어로 이어졌다. 그는 1909년 벨기에 루뱅 대학교의 교수 프란스 알폰스 얀센스가 발견하고 '키아스마티피'(chiasmatypiefra)라고 명명한 현상, 즉 염색체 간의 교차 가능성에 주목했다.[17] 모건은 연관된 유전자 사이의 교차 빈도가 다르며, 이 빈도가 염색체 상에서 유전자들을 분리하는 거리를 나타낼 수 있다고 제안했다. 즉, 교차 빈도가 높을수록 유전자 사이의 거리가 멀다는 것이다.
이러한 개념을 바탕으로 모건의 제자 앨프리드 스터티번트는 1913년 초파리의 X염색체에 대한 최초의 유전자 지도를 작성했다.[18] 이 지도는 백색 눈(white), 황색 몸(yellow), 절단 날개(miniature), 주홍색 눈(vermilion) 등 6개의 성 연관 유전자의 상대적 위치와 거리를 표시한 것이었다. 이는 유전자가 염색체 상에 실제로 존재하며 특정 위치를 차지한다는 염색체설의 강력한 증거가 되었다.
모건은 컬럼비아 대학교의 동료 연구자들인 스터티번트, 캘빈 브리지스, H. J. 멀러와 함께 초파리 연구를 포괄적인 유전 이론으로 발전시켰다. 1915년 이들은 연구 초기 결과를 요약한 《멘델 유전의 메커니즘》(The Mechanism of Mendelian Heredity)이라는 책을 출판했다.[20] 이 책에서 그들은 유전자가 염색체 지도 상의 특정 위치에 놓일 수 있음을 명확히 제시했으며, 세포학적 연구를 통해 지도상의 위치가 염색체상의 실제 위치와 일치함을 증명하여 멘델의 유전 요소가 염색체 구조에 물리적 토대를 두고 있음을 결정적으로 입증했다. 유전학자 커트 스턴(Curt Stern)은 이 책을 "새로운 유전학의 기본 교과서"라고 평가했다.[21]
모건 연구팀의 염색체 지도 작성은 유전학 연구의 새로운 장을 열었으며, 유전자가 추상적인 개념이 아니라 염색체 위에 존재하는 물리적 실체임을 증명하는 결정적인 계기가 되었다. 이후 유전적 연관성을 측정하는 단위는 영국의 유전학자 J. B. S. 홀데인에 의해 모건의 이름을 따 센티모건(centimorgan, cM)으로 제안되었다.
4. 4. 멘델 유전의 메커니즘
1908년부터 초파리(''Drosophila melanogaster'')를 이용한 유전 연구를 시작했다. 당시 모건은 다윈의 자연 선택 이론에 회의적이었으며, 휴고 드 브리스가 제안한 돌연변이에 의한 종 형성 이론을 실험적으로 증명하고자 했다. 그는 또한 멘델의 유전법칙과 염색체설에 대해서도 처음에는 회의적인 입장이었다.[15]모건은 페르난두스 페인(Fernandus Payne)과 함께 물리적, 화학적, 방사선적 방법으로 초파리의 인위적인 돌연변이를 유도하려 했으나[17][16], 2년간 뚜렷한 성과를 얻지 못했다.[17] 마침내 1910년, 그는 자신이 기르던 붉은 눈의 야생형 초파리 집단에서 흰 눈을 가진 돌연변이 수컷 한 마리를 우연히 발견했다. 이 흰 눈 수컷을 정상적인 붉은 눈 암컷과 교배시키자, 제1대 자손(F1)은 모두 붉은 눈을 가졌다. 다시 F1 세대끼리 교배하여 얻은 제2대 자손(F2)에서는 붉은 눈과 흰 눈 초파리가 나타났는데, 놀랍게도 흰 눈을 가진 초파리는 모두 수컷이었다.
이 결과를 설명하기 위해 모건은 이 형질이 성 염색체와 연관되어 유전된다는, 즉 반성 유전 가설을 세웠다. 그는 흰 눈 형질이 암컷의 X염색체와 관련이 있을 것으로 추정하고, 이 유전자를 '백색(white)'이라고 명명했다. 이후 다른 여러 돌연변이 형질들도 반성 유전된다는 사실을 발견하면서, 모건은 X염색체 위에 여러 유전자가 존재한다고 확신하게 되었다. 그는 1909년 덴마크의 식물학자 빌헬름 요한센이 도입한 유전자 개념을 받아들여, 유전자들이 염색체 위에 일직선으로 배열되어 있을 것이라고 결론지었다.
모건은 그의 제자들인 앨프리드 스터티번트, 캘빈 브리지스, 헤르만 조지프 멀러 등과 함께 초파리 연구를 더욱 발전시켰다. 특히, 성 염색체에 위치하면서도 때때로 독립적으로 유전되는 것처럼 보이는 소형 날개 돌연변이 등을 관찰하면서 유전자 연관과 염색체 교차 개념을 발전시켰다. 그는 1909년 벨기에의 프란스 알폰스 얀센스가 발견하고 '키아스마티피'라고 명명한 현상에 주목하여, 연관된 유전자들 사이에서 교차가 일어나며, 그 빈도가 유전자 사이의 거리를 나타낼 수 있다고 제안했다. 이를 바탕으로 그의 제자 앨프리드 스터티번트는 1913년 세계 최초의 유전자 지도를 작성했다.[18] 이는 유전자가 염색체 상의 특정 위치에 존재한다는 강력한 증거가 되었다.
1915년, 모건, 스터티번트, 브리지스, 멀러는 그들의 연구 결과를 집대성한 기념비적인 저서 《The Mechanism of Mendelian Heredity|멘델 유전의 메커니즘eng》을 출판했다.[20] 이 책은 현대 유전학의 기초를 다진 교과서로 평가받는다.[21] 이러한 연구들을 통해 모건은 초기에 가졌던 멘델 유전법칙과 염색체설에 대한 회의적인 입장을 버리고, 이 두 이론이 상호 보완적으로 유전 현상을 설명할 수 있음을 받아들였다. 그의 연구는 1902년~1903년 월터 서튼과 테오도어 보베리가 독립적으로 제안했던 멘델-염색체설을 실험적으로 증명하고 확립하는 데 결정적인 기여를 했다.
모건의 연구는 유전자를 추상적인 개념이 아닌, 염색체 위에 위치하는 물리적 실체로 인식하게 만들었으며, 유전학 연구의 새로운 길을 열었다. 그의 연구실, 이른바 '플라이룸(Fly Room)'은 세계적인 유전학 연구의 중심지가 되었고, 초파리는 유전학 연구의 가장 중요한 모델 생물 중 하나로 자리 잡게 되었다.[24]
비록 멘델-염색체설은 점차 학계의 인정을 받았지만, 유전자의 정확한 본질이나 작용 방식에 대해서는 여전히 논쟁이 있었다. W. E. 캐슬과 같은 학자들은 유전자 간의 상호작용을 강조했고, 리차드 골드슈미트 등은 유전자가 염색체 상의 불연속적인 단위라는 생각에 의문을 제기했다.[23]
모건은 1915년 이후 성장하던 우생학 운동과 그것이 인종 차별적 견해를 정당화하는 방식에 대해 강력히 비판하는 입장을 취했다.[26] 1928년, 그는 캘리포니아 공과대학교로 자리를 옮겨 생물학과를 창설하는 데 주도적인 역할을 했으며, 이후에도 염색체 유전설 연구를 계속하는 한편, 초기 관심사였던 실험발생학 연구로 돌아가 유전학과 발생학의 연결 고리를 찾으려 노력했다.
5. 발생학과 진화론 연구
1893년 모건은 발생학의 근본적인 문제 해결을 위해 실험적 방법을 도입했다. 그는 초기 배아 세포인 할구가 어떻게 배로 발달하는지, 유핵난세포와 무핵난세포의 수정 문제 등을 실험적으로 검토하며 발생 과정의 인과 관계를 밝히고자 했다. 1904년에는 컬럼비아 대학교의 실험동물학 교수로 부임하여 이후 24년간 중요한 유전 연구의 기반을 닦았다.
19세기 말 당시 많은 생물학자들처럼 모건 역시 초기에는 다윈의 진화론, 특히 자연선택 메커니즘에 대해 회의적이었다. 그는 복잡한 생물 적응 현상이 우연한 변이의 축적으로 설명될 수 있는지 의문을 가졌고, 다윈이 변이의 기원과 유전 메커니즘을 명확히 제시하지 못했다고 보았다. 진화 자체는 사실로 인정했지만, 실험적 검증이 부족한 자연선택은 불완전한 이론으로 여겼다. 마찬가지로 멘델의 이론과 염색체 이론에 대해서도, 세포 내에 특정 유전 단위가 미리 존재한다고 가정하는 점이 과거의 전성설과 유사하다며 비판적인 입장을 취했다. 그는 염색체와 유전 현상의 관련성은 인정했지만, 1909년 무렵까지 개별 염색체가 특정 형질을 결정한다는 생각에는 동의하지 않았고, 멘델의 법칙 역시 유전의 구체적인 과정을 설명하지 못하는 가설적 이론으로 간주했다.
1928년 모건은 캘리포니아 공과대학교(Caltech)로 옮겨 생물학부 부서장을 맡았고, 1942년 은퇴할 때까지 14년간 재직했다. 그는 칼텍 생물학부를 유전학과 진화, 실험 발생학, 생리학, 생물 물리학, 생화학 연구 중심지로 만들고자 했으며, 코로나 델 마르에 해양 연구소 설립에도 기여했다. 브리지스, 스터트번트, 테오도시우스 도브잔스키, 조지 비들, 라이너스 폴링 등 당대 최고의 과학자들을 영입하여 연구 역량을 강화했다.
모건은 학계에서도 중요한 역할을 수행했다. 1927년부터 1931년까지 미국 과학 아카데미 회장을, 1930년에는 미국 과학 진흥 협회 회장을 역임했으며, 1932년 제6회 국제 유전학 회의 의장을 맡았다. 1933년에는 유전 연구의 공로를 인정받아 노벨 생리학·의학상을 수상했다. 그는 이 성과가 공동 연구의 결과임을 강조하며 상금을 동료 연구자들과 자녀들에게 나누어 주었다. 같은 해 초파리 침샘에서 거대 다사 염색체가 발견되면서, 이전까지 표현형 관찰을 통해 추론했던 유전 연구 결과들을 물리적 증거로 확인할 수 있게 되었다. 1939년에는 영국 왕립학회로부터 코플리 메달을 받았다.
초파리 연구를 통해 유전 가능한 돌연변이를 다수 발견하면서 모건의 생각은 점차 변화했다. 유전 메커니즘에 대한 이해가 깊어지자 그는 자연선택 이론을 새로운 관점에서 검토하게 되었다. 1916년 저서 ''A Critique of the Theory of Evolution|진화론 비판eng''에서 그는 멘델 유전학과 염색체 이론을 바탕으로 진화가 생존에 유리한 돌연변이가 집단 내에 통합되는 과정임을 설명했다.[31][32] 비록 초기에는 다윈주의를 비판했지만, 그의 연구는 유전학의 기초를 확립하고 자연선택 이론의 핵심 기반을 제공함으로써 신다윈주의 종합에 크게 기여했다.
모건은 평생 진화에 깊은 관심을 가졌으며, 관련 논문과 책을 저술했다.[27] 1942년 은퇴 후에도 명예 교수로서 성 분화, 재생, 발생학 등 진화와 관련된 연구를 이어갔다.
5. 1. 실험 발생학 연구
1890년, 모건은 존스 홉킨스 대학교의 자매 학교인 브린모어 대학교(Bryn Mawr College)의 부교수(겸 생물학과장)로 임명되어, 동료 에드먼드 비처 윌슨(Edmund Beecher Wilson)의 뒤를 이었다.[7] 그는 형태학 관련 과목을 가르쳤고, 자크 뢰브(Jacques Loeb)는 생리학 과목을 담당했다.[8] 초기 몇 년간 모건은 바다 따개비, 멍게 벌레, 개구리에 대한 기술적 연구를 수행했다.1893년부터 모건은 발생학의 근본적인 문제를 해결하기 위해 실험적 방법을 도입했다. 그는 발생 과정의 인과 관계를 밝히고자 초기 배아 세포인 할구(blastomere)가 어떻게 완전한 배로 발달하는지, 유핵난세포와 무핵난세포의 수정 문제 등을 실험적으로 검토했다.
1894년에는 1년간의 연구 휴가를 얻어 나폴리(Naples)의 ''Stazione Zoologica'' 해양 연구소에서 연구했다. 이곳에서 독일 생물학자 한스 드리쉬(Hans Driesch)와 함께 연구하며 그의 실험 발생학 연구에 큰 영향을 받았다. 모건은 이 시기 빗해파리 발생학에 대한 실험 연구를 수행했으며, 뢰브를 통해 생명 현상을 물리적, 화학적으로 설명하려는 독일의 ''Entwicklungsmechanik'' (발달 역학) 학파를 접하게 되었다. 이는 모건이 전통적인 기술(記述) 중심의 형태학에서 실험 발생학으로 연구 방향을 전환하는 중요한 계기가 되었다.[9]
당시 발생학계에서는 배아 발달 과정을 둘러싼 논쟁이 활발했다. 빌헬름 루(Wilhelm Roux) 등은 유전 물질이 배아 세포마다 나뉘어 각 세포가 특정 기관으로 발달하도록 미리 정해져 있다는 '모자이크 이론'을 주장했다. 반면, 드리쉬와 모건은 난자의 세포질과 핵, 그리고 환경 사이의 상호작용이 발달에 영향을 미친다는 '후성적 발달론'을 지지했다. 모건은 드리쉬와의 공동 연구를 통해 성게와 빗해파리 난자에서 분리된 초기 세포(세포분열구)가 각각 완전한 유충으로 발달할 수 있음을 실험적으로 증명하며, 모자이크 이론의 예측과 달리 개별 세포가 전체 개체를 형성할 잠재력을 가지고 있음을 보였다.[10] 또한, 발생 과정에서 후성유전 및 환경 요인의 중요성을 탐구하는 과정에서, 성게 난자에 염화마그네슘을 처리하여 수정 없이도 세포 분열을 유도할 수 있음을 보여주었다. 이는 후에 뢰브가 인공적인 단위생식 연구로 발전시키는 계기가 되었다.[11][12]
1895년 브린모어 대학교로 돌아와 정교수로 승진한 모건은 재생과 유충 발달에 대한 실험 연구를 계속하며 발생의 내적 요인과 외적 요인을 구분하고자 했다. 그는 1897년 첫 저서인 ''개구리 난자의 발달''(The Development of the Frog's Egg)을 출판했고, 올챙이, 물고기, 지렁이 등 다양한 유기체의 재생 능력을 연구하여 1901년 ''재생''(Regeneration)이라는 책으로 정리했다.
19세기 말의 많은 생물학자들처럼 모건 역시 초기에는 자연선택을 통한 다윈의 진화론에 회의적이었다. 복잡한 생명 현상이 우연한 변이의 축적으로 설명될 수 있는지 의문을 가졌고, 다윈이 변이의 기원과 유전 메커니즘(판게네시스 가설 제외)을 명확히 제시하지 못했다고 보았다. 그는 진화 자체는 사실로 받아들였지만, 실험적 검증이 부족한 자연선택 메커니즘은 불완전하다고 생각했다.
마찬가지로 그는 멘델의 이론과 염색체 이론에 대해서도 비판적인 입장이었다. 이 이론들이 세포 내에 특정 유전 단위가 존재한다고 가정하는 것이 마치 과거의 전성설(preformationism)과 유사하다고 보았기 때문이다. 그는 염색체가 유전 현상과 관련이 있다는 점은 인정했지만, 1909년 무렵까지도 개별 염색체가 특정 유전 형질을 결정한다는 생각에는 동의하지 않았다. 멘델의 법칙이 교배 결과를 설명하고 예측할 수는 있지만, 유전의 구체적인 과정을 증명하지는 못하는 순전히 가설적인 이론이라고 주장했다. 염색체 쌍의 분리와 멘델의 유전 인자 분리가 일치한다는 정황만으로는 두 현상의 직접적인 연관성을 확신하기 어렵다고 보았다.
1900년부터는 성 결정 시스템 문제에 대한 연구를 시작했으며, 이는 네티 스티븐스(Nettie Stevens)가 Y 염색체의 역할을 발견한 이후 더욱 본격화되었다. 그는 발생학 연구와 더불어 진화 문제에 대한 탐구도 계속 이어나갔다.[13]
5. 2. 진화론 연구와 유전학의 통합
19세기 말, 당시 많은 발생학자나 생물학자들처럼 모건 역시 다윈의 자연선택설이 생물체의 복잡한 적응 현상을 설명하기에는 설득력이 부족하다고 생각했다. 사소하고 우연적인 변이가 축적되어 복잡한 적응이 이루어진다는 개념을 받아들이기 어려웠으며, 다윈이 변이의 기원과 유전 메커니즘을 명확히 제시하지 못했다는 점(초기 가설인 판게네시스 제외)을 비판했다. 모건은 진화 자체는 사실로 받아들였지만, 실험적으로 검증할 수 없는 자연선택 메커니즘은 불완전한 이론으로 간주했다.그는 멘델의 이론과 염색체설에 대해서도 비판적이었다. 이 이론들이 세포 내에 발생을 조절하는 유전적 단위나 물질적 실체가 있다고 가정하는 방식이, 과거 전성설(미리 형성된 개체가 생식세포 안에 축소되어 있다는 학설)과 유사하다고 보았다. 1909년부터 약 1년간 그는 염색체가 유전 현상과 관련 있다는 점은 인정하면서도, 개별 염색체가 특정한 유전 형질을 직접 결정하지는 않는다고 주장했다. 또한 멘델의 이론이 교배 결과를 설명하고 예측할 수는 있지만, 유전의 정확한 과정을 설명하지 못하므로 순전히 가설적이라고 보았다. 염색체 쌍의 분리와 각 염색체가 서로 다른 생식세포로 들어간다는 사실만으로는 두 과정(멘델의 유전과 염색체 행동)이 직접 관련 있다는 증거가 부족하다고 생각했다.
1903년 저서 ''Evolution and Adaptation|진화와 적응eng''에서 그는 미세한 개체 차이에 작용하는 선택만으로는 완전히 새로운 종을 만들 수 없다는 반(反)다윈주의적 입장을 명확히 했다.[27] 그는 다윈의 성 선택 이론[28]과 획득 형질의 유전이라는 신라마르크주의 이론[29] 역시 거부했다. 당시(1875년~1925년)는 '다윈주의의 쇠퇴'라고 불릴 정도로 자연선택에 대한 비판이 많았던 시기였으며, 모건도 그 비판자 중 한 명이었다.[30]
그러나 초파리 연구를 통해 작고 안정적이며 유전되는 돌연변이를 다수 발견하면서 모건의 생각은 점차 바뀌기 시작했다. 돌연변이가 유전된다는 사실은 진화에 직접적인 영향을 미칠 수 있음을 의미했기 때문이다. 유전 문제에 대한 이해가 깊어지면서(대략 1915년 경), 그는 다윈의 자연선택 이론을 새로운 관점에서 비판적으로 검토할 수 있게 되었다.
1916년 출간된 ''A Critique of the Theory of Evolution|진화론 비판eng''에서 모건은 "선택은 진화에서 어떤 역할을 하는가?", "선택은 어떻게 새로운 것을 만들어낼 수 있는가?" 등의 질문을 던지며 진화 메커니즘을 탐구했다. 이 책에서 그는 멘델 유전학과 그 염색체적 기반을 상세히 설명하며, "야생 동식물과 가축 품종의 형질 모두 멘델의 법칙에 따라 유전된다"고 결론지었다. 그는 진화가 유기체의 생존과 번식에 유리한 돌연변이가 집단 내에 통합됨으로써 일어난다고 보았으며, 해로운 돌연변이는 자연선택에 의해 제거되어 정착되기 어렵다고 설명했다.[31][32]
결국 모건은 진화 자체를 부정한 것이 아니라, 오히려 진화의 핵심 메커니즘인 유전의 기초를 확립했다. 다윈은 자연선택 이론을 제시했지만 유전의 작동 원리를 설명하지 못했는데, 모건은 바로 이 부분을 해결함으로써 자연선택 이론의 과학적 토대를 마련한 것이다. 비록 경력 초기에는 다윈주의를 비판했지만, 그의 연구는 결과적으로 신다윈주의 종합(유전학과 진화론의 통합)에 크게 기여했다.
모건은 평생 진화에 깊은 관심을 가졌으며, 해양거미류( sea spiderseng )의 계통 발생에 관한 논문을 쓰고 진화에 관한 네 권의 책을 저술했다.[27] 1933년 노벨 생리학·의학상 수상 연설에서는 유전 상담 외에 유전학이 의학에 기여할 수 있는 부분을 언급하기도 했다. 은퇴 후에도 그는 성 분화, 재생, 발생학 등 진화와 관련된 연구를 계속했다.
6. 수상 및 영예
1909년 미국 국립 과학 아카데미 회원으로 선출되었고,[34] 1915년에는 미국 철학 학회 회원,[35] 1919년에는 왕립 학회 외국인 회원(ForMemRS)으로 선출되었다.[1] 1928년에는 미국 예술 과학 아카데미 회원으로도 선출되었다.[36]
모건은 여러 학술 단체에서 중요한 역할을 수행했다. 1927년부터 1931년까지 미국 과학 아카데미 회장을 지냈으며, 1930년에는 미국 과학진흥협회 회장, 1932년에는 제6차 국제유전학회 회장으로 활동했다.
그는 유전학 분야에서의 뛰어난 업적을 인정받아 다음과 같은 주요 상들을 수상했다.
또한 존스 홉킨스 대학교에서 명예 법학박사(LL.D.) 학위를, 켄터키 대학교에서 명예 철학박사(Ph.D.) 학위를 받았다.
모건의 이름은 그의 업적을 기리기 위해 여러 기관과 상에 사용되고 있다.
- 켄터키 대학교의 토머스 헌트 모건 생물 과학 학교는 그의 이름을 따서 명명되었다.
- 미국 유전학회는 매년 유전학 분야에 큰 기여를 한 회원에게 그의 이름을 딴 토머스 헌트 모건 메달을 수여한다.
- 1989년 스웨덴에서 발행된 우표에는 그의 발견이 다른 8명의 노벨상 수상 유전학자들의 발견과 함께 묘사되었다.
- 워싱턴 주 쇼어라인에 있는 한 중학교는 20세기 후반에 모건의 이름을 따서 명명되었다.
모건은 유전학 분야에 중요한 유산을 남겼으며, 그의 제자들 중 조지 웰스 비들과 헤르만 조셉 뮬러를 포함한 여러 명이 노벨상을 수상했다. 노벨상 수상자인 에릭 캔들은 모건의 유전자와 염색체 위치에 대한 발견이 생물학을 실험 과학으로 전환하는 데 크게 기여했다고 평가했다.[33]
7. 유산과 영향
모건은 유전학 분야에 중요한 유산을 남겼다. 컬럼비아 대학교와 캘리포니아 공과대학교에서 배출된 그의 제자들 중에는 노벨상 수상자인 조지 웰스 비들과 헤르만 조셉 뮬러 등이 있다. 노벨상 수상자 에릭 캔들은 모건에 대해 "다윈의 동물 종 진화에 대한 통찰이 19세기 생물학에 설명을 더하는 과학으로서 처음으로 일관성을 부여한 것처럼, 모건의 유전자와 염색체 위치에 대한 발견은 생물학을 실험 과학으로 바꾸는 데 기여했다"고 평가했다.[33]
모건은 다양한 학술 기관으로부터 인정을 받았으며, 그의 업적을 기리는 여러 활동이 이루어졌다.
'''주요 수상 및 영예'''
| 연도 | 내용 |
|---|---|
| 1909년 | 미국 국립 과학 아카데미 회원 선출[34] |
| 1915년 | 미국 철학 학회 회원 선출[35] |
| 1919년 | 왕립 학회 외국인 회원(ForMemRS) 선출[1] |
| 1924년 | 다윈 메달 수상 |
| 1928년 | 미국 예술 과학 아카데미 회원 선출[36] |
| 연도 미상 | 존스 홉킨스 대학교 명예 법학박사(LL.D.) 학위 수여 |
| 연도 미상 | 켄터키 대학교 명예 철학박사(Ph.D.) 학위 수여 |
'''기념'''
- 켄터키 대학교의 토머스 헌트 모건 생물 과학 학교는 그의 이름을 따서 명명되었다.
- 미국 유전학회는 유전학 발전에 크게 기여한 회원에게 매년 그의 이름을 딴 토머스 헌트 모건 메달을 수여한다.
- 1989년 스웨덴에서 발행된 우표에는 모건을 포함한 8명의 노벨상 수상 유전학자들의 발견이 묘사되었다.
- 미국 워싱턴 주 쇼어라인에 위치한 한 중학교는 20세기 후반에 모건의 이름을 따서 명명되었다.
8. 개인적 삶
1904년 6월 4일, 모건은 브린마르 대학교에서 생물학 대학원에 재학 중이던 릴리안 본 섐슨(1870년–1952년)과 결혼했다. 그녀는 결혼 후 16년간 과학 연구를 중단하고 네 명의 자녀(아들 1명, 딸 3명)를 양육했다. 이후 릴리안은 모건의 초파리 연구에 크게 기여했다. 네 자녀 중 한 명인 이서벨 모건(1911년–1996년)은 소아마비 연구를 전문으로 하는 바이러스학자가 되었다. 모건은 무신론자였다.
참조
[1]
논문
Thomas Hunt Morgan. 1866–1945
[2]
웹사이트
The Nobel Prize in Physiology or Medicine 1933
http://nobelprize.or[...]
Nobel Web AB
2010-09-14
[3]
문서
Sturtevant (1959), p. 283.
[4]
문서
Allen (1978), pp. 11–14, 24.
[5]
문서
Allen, ''Thomas Hunt Morgan: The Man and His Science'', pp. 46–51
[6]
논문
Thomas Hunt Morgan at the Marine Biological Laboratory: Naturalist and Experimentalist
[7]
논문
Edmund Beecher Wilson. 1856–1939
[8]
문서
Allen, ''Thomas Hunt Morgan'', pp. 50–53
[9]
문서
Allen, ''Thomas Hunt Morgan'', pp. 55–59, 72–80
[10]
문서
Allen, ''Thomas Hunt Morgan'', pp. 55–59, 80–82
[11]
논문
On the Nature of the Process of Fertilization and the Artificial Production of Normal Larvae (Plutei) from the Unfertilized Eggs of the Sea Urchin
http://www.stanford.[...]
[12]
서적
Artificial parthenogenesis and fertilization
https://archive.org/[...]
University of Chicago Press
[13]
문서
Allen, ''Thomas Hunt Morgan'', pp. 84–96
[14]
문서
Allen, ''Thomas Hunt Morgan'', pp. 68–70
[15]
문서
Allen, ''Thomas Hunt Morgan: The Man and His Science'', pp. 105–116
[16]
논문
The Secrets of Life: Historian Luis Campos resurrects radium's role in early genetics research
https://www.scienceh[...]
2018-03-22
[17]
문서
Kohler, ''Lords of the Fly'', pp. 37–43
[18]
논문
The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association
http://www.esp.org/f[...]
1913
[19]
서적
Biology Ninth Edition
McGraw-Hill
2007
[20]
서적
The Mechanism of Mendelian Heredity
https://books.google[...]
Henry Holt
[21]
논문
The Continuity of Genetics
https://www.jstor.or[...]
1970
[22]
서적
The theory of the gene
http://archive.org/d[...]
New Haven, Yale University Press; [etc., etc.]
1926
[23]
문서
Allen, ''Thomas Hunt Morgan'', pp. 208–213, 257–278. Quotation from p. 213.
[24]
문서
Kohler, ''Lords of the Fly'', chapter 5
[25]
문서
Allen, ''Thomas Hunt Morgan'', pp. 214–215, 285
[26]
문서
Allen, ''Thomas Hunt Morgan'', pp. 227–234
[27]
서적
Evolution. The First Four Billion Years
https://archive.org/[...]
Harvard University Press
[28]
문서
"I think we shall be justified in rejecting it as an explanation of the secondary sexual differences amongst animals", pp. 220–221, chapter VI, ''Evolution and Adaptation'', 1903.
[29]
문서
Chapter VII of ''Evolution and Adaptation'', 1903.
[30]
서적
Evolution. The History of an Idea
University of California Press
[31]
서적
'' A Critique of the Theory of Evolution'', Princeton University Press, 1916, pp. 193–194
[32]
문서
''A Critique of the Theory of Evolution'', p. 189.
[33]
웹사이트
Genes, Chromosomes, and the Origins of Modern Biology
http://www.columbia.[...]
1999
[34]
웹사이트
Thomas Morgan
http://www.nasonline[...]
2019-04-28
[35]
웹사이트
APS Member History
https://search.amphi[...]
2023-11-02
[36]
웹사이트
Thomas Hunt Morgan
https://www.amacad.o[...]
2023-11-02
[37]
서적
Strange Angel: The Otherworldly Life of Rocket Scientist John Whiteside Parsons
Houghton Mifflin Harcourt
[38]
웹사이트
T. H. Morgan at Caltech: A Reminiscence
http://www.genetics.[...]
1998-08-01
[39]
웹사이트
The Thomas Hunt Morgan Era in Biology
https://web.archive.[...]
2019-04-28
[40]
학술지
T. H. Morgan at Caltech: A Reminiscence
https://web.archive.[...]
1998-08-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com